金币
UID746503
帖子
主题
积分1492
注册时间2021-9-24
最后登录1970-1-1
听众
性别保密
|
欢迎您注册蒲公英
您需要 登录 才可以下载或查看,没有帐号?立即注册
x
肿瘤来自基因突变,这是人体衰败的主因,也是人类进化的根源。人体内外因素一直驱动着肿瘤基因组发生体细胞突变,并在这一过程中形成了具有特征性的突变标签集。我们今天的主角 APOBEC3就与癌症中一些最普遍的突变特征有关。
载脂蛋白 B mRNA 编辑酶催化多肽 (apolioprotein B mRNA-editing enzyme catalytic polypeptide, APOBEC) 家族是一类高效的胞苷脱氨酶,能够特异催化基因组中的胞嘧啶变为尿嘧啶(C>U),其转录可被促炎症细胞因子和趋化因子激活,并在人体的固有和获得性免疫机制中发挥重要作用,在慢性炎症条件下,APOBEC 家族表达异常,会误伤人类基因组引发 APOBEC 突变特征。
早期对肿瘤基因组突变模式的研究表明,胞嘧啶突变通常存在于 TCN(N代表ATCG中任意核苷酸)序列部分,而通过来自肿瘤基因组的单碱基替换(single-base substitutions,以下简称SBS)可显示出 APOBEC 相关胞嘧啶突变的两类不同突变特征:非聚集型(SBS2 和 SBS13)和聚集型(kataegis 和 omikli)。
目前的研究表明,已在 超过 70% 的肿瘤类型和约 50% 的肿瘤基因组中发现了 APOBEC 相关的突变特征,特别是在乳腺癌和膀胱癌中尤为突出。但是,内源性 APOBEC3 与人类癌症基因组突变特征之间的关系尚不明晰。
近日,美国 MIT 和哈佛大学的 Mia Petljak 等人在 Nature 上发表了文章 Mechanisms of APOBEC3 mutagenesis in human cancer cells,建立了二者之间的联系。
 [backcolor=rgba(18, 18, 18, 0.5)]​ [backcolor=rgba(18, 18, 18, 0.5)]​
编辑切换为居中
添加图片注释,不超过 140 字(可选)
肿瘤细胞中 APOBEC 突变
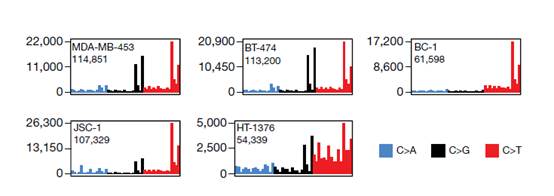
为了研究 APOBEC3 诱变的机制,作者从一系列癌细胞系中分别敲除了 APOBEC3A 和 APOBEC3B 基因,发现这些癌细胞系随着时间的推移都产生了 APOBEC3 相关突变,并且 APOBEC3B 是靶向线性和发夹结构的胞嘧啶脱氨酶的主要来源。
 [backcolor=rgba(18, 18, 18, 0.5)]​ [backcolor=rgba(18, 18, 18, 0.5)]​
编辑
细胞系的突变以胞嘧啶碱基的全基因组替换数(y轴)表示,被划分为48种可能的三核苷酸序列
另外,作者还进一步分析了 APOBEC3 相关的突变特征。对来自 251 个乳腺癌、膀胱癌和淋巴瘤癌细胞系克隆的全基因组序列突变特征的分析表明,APOBEC3A 的敲除减少了 APOBEC3 相关的 SBS2 和 SBS13a/b 突变特征。
与之相反,尽管 APOBEC3B 在所有细胞系中与 APOBEC3A 相比具有更高的表达和脱氨酶活性,但APOBEC3B 的敲除并没有显著减少 SBS2 和 SBS13a/b 类型突变,反而增加了 APOBEC3A 蛋白水平、活性和 APOBEC3A 介导的诱变。
另外,作者还通过研究肿瘤细胞中 APOBEC3 对 kataegis(聚焦链协同超突变)和 omikli(弥漫性超突变)的影响最终确定了APOBEC3A 是突变的主要驱动因素,这些数据都表明了内源性 APOBEC3 脱氨酶参与人类癌细胞的突变。
APOBEC3 突变机制
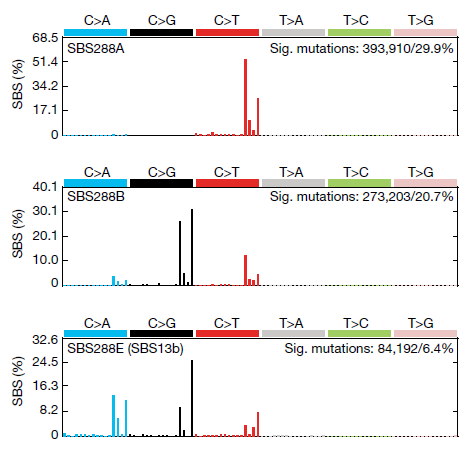
APOBEC3 相关突变主要为 SBS2 和 SBS13 a/b。SBS2 的特征是 C>T 突变,而 SBS13 的特征是 C>G 和 C>A 突变,APOBEC3 生成的尿嘧啶的加工过程可能决定了产生的突变类型。而作者认为 尿嘧啶的复制可能导致 C>T 突变,从而导致 SBS2。
 [backcolor=rgba(18, 18, 18, 0.5)]​ [backcolor=rgba(18, 18, 18, 0.5)]​
编辑
从 SBSs 中提取的 APOBECs 相关突变特征片段
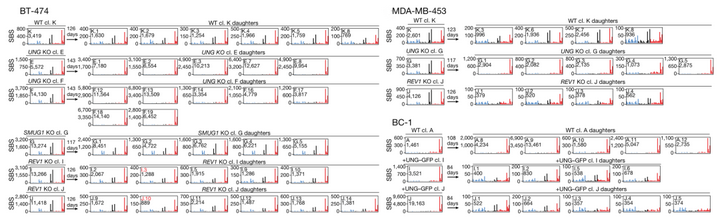
而通过糖基化酶(如UNG或SMUG1)切除尿嘧啶,然后再进行 TLS(translesion DNA synthesis),可能导致 C>T、C>G 和 C>A突变,从而产生 SBS 2 和 SBS 13 的组合。为了直接评估 UNG 和 SMUG1 对癌细胞中 SBS2 和 SBS 13a/b 产生的影响,研究人员在 BC-1 细胞中表达 UNG-GFP、在 BT-474 细胞中 CRISPR-Cas9 删除SMUG1、在 BT-474 和 MDA-MB-453 细胞中删除 UNG。
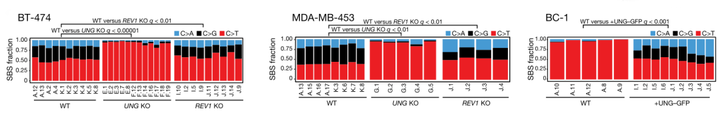
结果表明在 BRCA 细胞系子代克隆中,UNG 的缺失降低了 TCN 环境中 C>A 和 C>G 突变的相对比例,而 BC-1 细胞中 GFP-UNG 的表达增加了 BC-1 子代克隆中这些突变类型的比例,SBS13a/b 突变减少,但未被消除。
 [backcolor=rgba(18, 18, 18, 0.5)]​ [backcolor=rgba(18, 18, 18, 0.5)]​
编辑切换为居中
添加图片注释,不超过 140 字(可选)
 [backcolor=rgba(18, 18, 18, 0.5)]​ [backcolor=rgba(18, 18, 18, 0.5)]​
编辑切换为居中
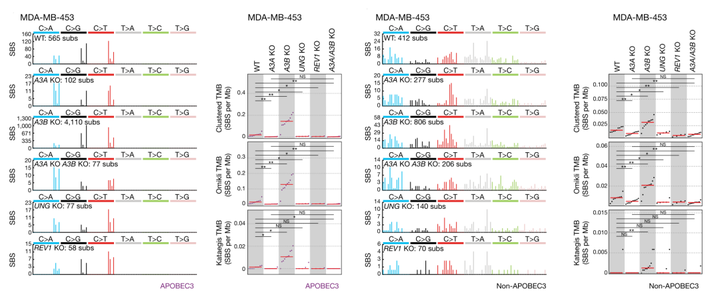
UNG 和 REV1 在癌症中 APOBEC3 突变的产生中起着关键作用
这些结果表明 UNG 将全基因组 APOBEC3 脱氨酶活性与转位突变联系了起来。UNG 的缺失导致 MDA-MB-453 细胞中 APOBEC3 相关突变和其他整体聚集突变的减少,但 UNG 活性与聚集突变的精确机制还需要进一步研究。
 [backcolor=rgba(18, 18, 18, 0.5)]​ [backcolor=rgba(18, 18, 18, 0.5)]​
编辑切换为居中
APOBEC3脱氨酶驱动人类癌细胞中聚集型突变的获得
而 SMUG1 缺失会导致 TCN 环境中 C>A 突变相对于 C>G 突变比例更高, SBS13b 增加。联系到在 UNG 敲除的子代克隆中观察到的持续 C>G/ A突变,以上这些结果表明 SMUG1 可能切除 APOBEC3 介导的尿嘧啶碱基, 且 SMUG1 可以代替 UNG 修复 U:G 基因组缺口。
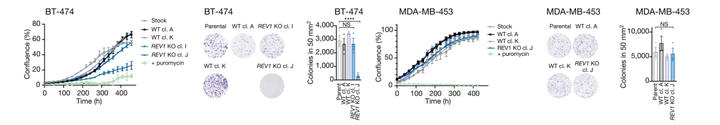
同时,为了评估 TLS 对 SBS 2 和 SBS13a/b 的影响,作者还在 BRCA 细胞系中构建了 REV1 特异性敲除细胞,发现 REV1 敲除子细胞中 SBS2 和 SBS13a/b 突变减少。
在 REV1 敲除子基因中 C>G 比例的降低可能反映了 REV1 脱氧胞苷转移酶活性的丧失,而 SBS2 和 SBS13a/b 的负担减少可能是由于 REV1 失去了作为其他 Y 家族聚合酶协调的支架的非催化作用。这些结果直接将REV1与人类癌细胞基因组中 APOBEC3 介导的突变特征的产生联系起来。除了 APOBEC3 相关突变外,在来自两个 BRCA 细胞系的 REV1 敲除子细胞中,发生在 APOBEC3 相关序列之外的其他 SBS 和聚集突变的负担分别减少。这些观察结果表明REV1 调节了各种 SBS 类型。
此外,REV1 敲除细胞的增殖、克隆存活、APOBEC3A 蛋白水平或 APOBEC3 催化活性下降并不一致,REV1 敲除细胞也没有表现出细胞周期动力学改变或 DNA 损伤增加。
 [backcolor=rgba(18, 18, 18, 0.5)]​ [backcolor=rgba(18, 18, 18, 0.5)]​
编辑切换为居中
REV1敲除细胞APOBEC3A蛋白水平或APOBEC3催化活性降低
而且对带有和不带有 APOBEC3 特征突变的细胞系进行的全基因组 Drop-out CRISPR 筛选分析未能显示在含有 SBS2/13 突变的癌细胞系中对 REV1 的依赖性增加。这些结果表明,REV1 在 SBS2 和 SBS13 的产生中发挥了关键作用,并且内源性 REV1 活性有助于基因突变。
总结
APOBEC3 家族的胞嘧啶脱氨酶与癌症中一些普遍的突变特征有关,但内源性 APOBEC3 与人类癌症基因组突变特征之间的因果关系尚未明确。
作者通过对人类肿瘤细胞的多重关联研究发现 APOBEC3A 的缺失是 APOBEC3 突变的主要驱动因素,其缺失可导致 APOBEC3 相关突变水平降低;而 APOBEC3B 的缺失会增加 APOBEC3A 的蛋白水平、活性以及 APOBEC3A 介导的突变。
此外,研究数据表明,UNG 和 REV1 依赖性 TLS 与人类肿瘤细胞中 APOBEC3 诱导的突变有关,尿嘧啶糖基化酶 UNG 是 APOBEC3 介导的转化所必需的,内源性 REV1 的缺失会降低细胞整体突变水平。
▎陶术生物内容团队编辑
版权说明:本文来自陶术生物内容团队,欢迎个人转发至朋友圈,谢绝媒体或机构未经授权以任何形式转载至其他平台。
如果您觉得内容不错,欢迎点赞、评论、分享给其他读者哦!关注公众号:陶术生物——药物筛选一站式平台,了解最新药筛研究资讯及促销活动或购买相关产品,如您有构建筛选化合物库的需求,欢迎通过以下方式联系我们,我们可以结合您的筛选需求及市场热点推荐最合适的化合物库构建方案。了解更多化合物库信息,也可点击陶术生物官网查询。
陶术只为有资质的机构和企业用于科学研究,不为任何个人,或者非科研性质的其他用途提供服务。
|
|

 |手机版|蒲公英|ouryao|蒲公英
( 京ICP备14042168号-1 ) 增值电信业务经营许可证编号:京B2-20243455 互联网药品信息服务资格证书编号:(京)-非经营性-2024-0033
|手机版|蒲公英|ouryao|蒲公英
( 京ICP备14042168号-1 ) 增值电信业务经营许可证编号:京B2-20243455 互联网药品信息服务资格证书编号:(京)-非经营性-2024-0033






 发表于 2022-8-16 10:36:04
发表于 2022-8-16 10:36:04

 置顶卡
置顶卡 变色卡
变色卡