金币
UID933789
帖子
主题
积分963
注册时间2024-1-9
最后登录1970-1-1
听众
性别保密
|

楼主 |
发表于 2024-2-1 16:25:45
|
显示全部楼层
肿瘤 “冷” 变 “热”?机制大盘点! - MedChemExpress
本帖最后由 18317190548 于 2024-2-1 16:27 编辑
肿瘤,始终是科研热榜的“TOP1”, 肿瘤除了有“良恶”之别,还有“冷热”之分。今天,咱们就一起来唠一唠肿瘤 “冷热” 机制那些事儿~

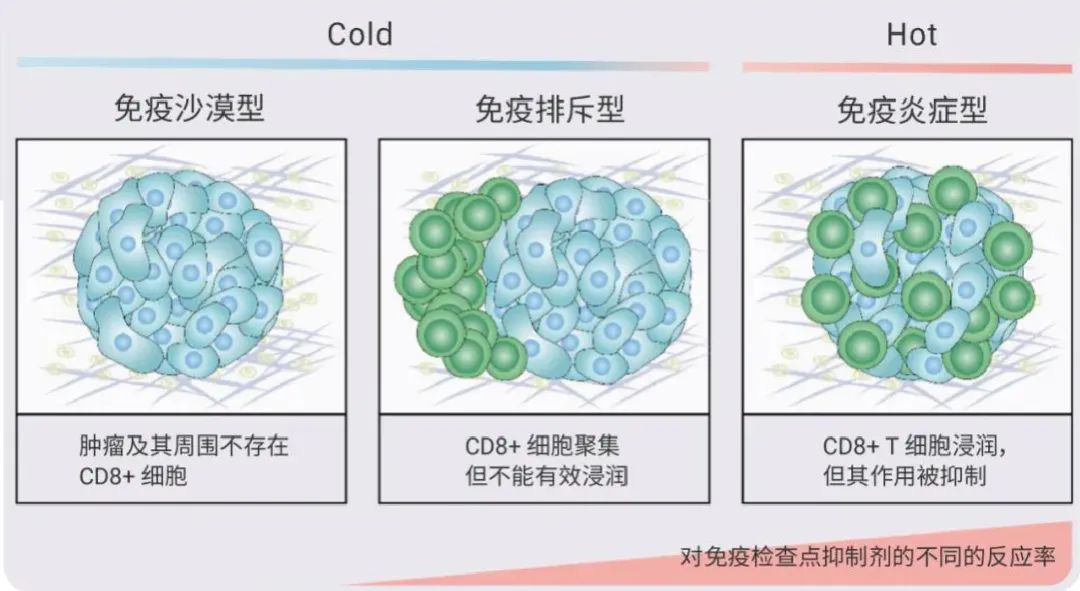
“冷热” 肿瘤如何划分?根据肿瘤微环境中免疫细胞的空间分布情况,将肿瘤分为三种基本的免疫表型:免疫炎症型、免疫排斥型和免疫沙漠型。其中免疫炎性肿瘤即为 “热肿瘤”,免疫排斥瘤和免疫沙漠瘤皆可称为 “冷肿瘤”。
简单来讲,“冷”肿瘤缺乏先天免疫,而在“热” 肿瘤中,免疫细胞较为活跃,其内环境也被大量的 T 细胞所浸润。
图 1. 根据免疫细胞分布区分“冷热”肿瘤[1]。
通常情况下,当免疫检查点抑制剂 (ICIs) 解除免疫检查点的抑制作用后,T 细胞对肿瘤的免疫应答效应再次被启动,活化的 T 细胞杀伤癌细胞,起到免疫治疗作用。
因此,具有炎症表型的 “热肿瘤” 往往对 ICIs 更敏感。但对于“冷肿瘤” 来说,免疫细胞很难识别并杀伤,免疫检查点抑制剂也就难以发挥作用,这也是导致“冷肿瘤”对 ICIs 治疗失效的主要原因[1]。
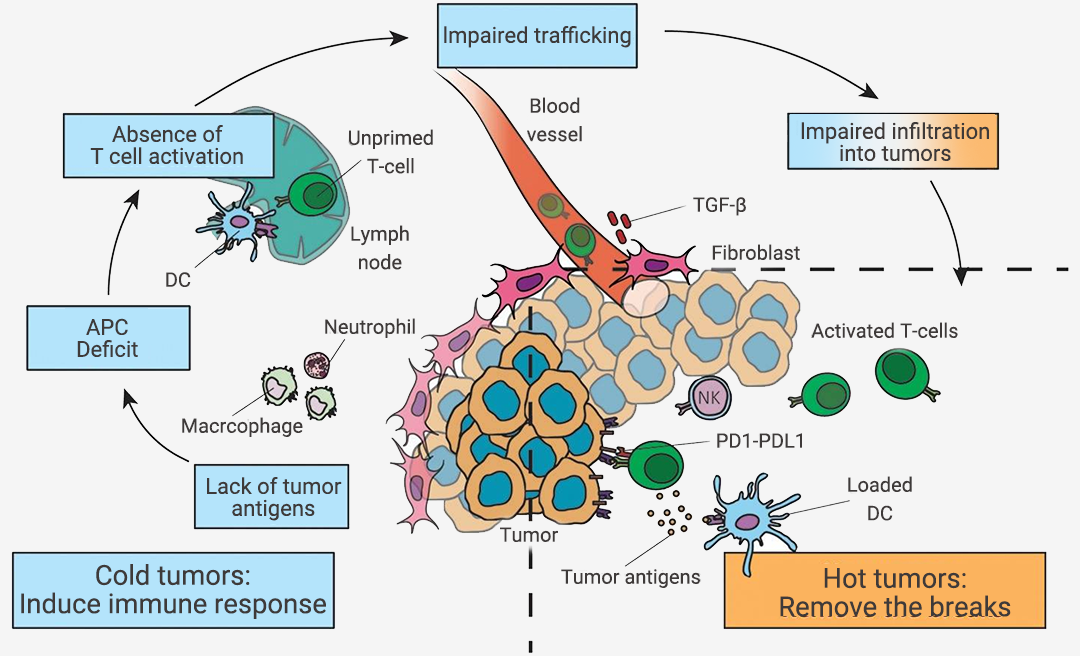
现有研究表明,使细胞毒性效应 T 细胞富集到“冷”肿瘤中,能够使“冷肿瘤”转变为“热肿瘤”从而显著提高肿瘤对免疫检查点抑制剂 ICIs 治疗的反应[2][3]。图 2. “冷”肿瘤转变为“热”肿瘤[4]。
在肿瘤免疫激活的过程中,冷肿瘤中 T 细胞缺失可能有以下原因[4]:
① 肿瘤相关抗原缺乏;
② 抗原呈递细胞 (APCs) 缺陷;
③ T 细胞活化缺失;
④ T 细胞向肿瘤的运输受损。
▐ 肿瘤相关抗原缺乏
人体的免疫系统能够识别健康细胞及肿瘤细胞,因为后者表达肿瘤相关抗原、肿瘤特异性抗原、癌症种系抗原,这些抗原会被体内的 CD8+ T 细胞识别并实现免疫应答[5]。目前研究表明,肿瘤抗原释放的动力学与不同形式的自发或诱导癌细胞死亡相关,最后导致 T 细胞的反应效果[6]。
▐ 抗原呈递细胞缺陷
肿瘤免疫在很大程度上依赖于 APCs 介导的效应 T 细胞的启动和激活,而树突细胞 (Dendritic cells, DCs) 是机体功能最强的 APCs。但在很多肿瘤中发现 DCs 是有缺陷的,造成这种缺陷的原因可能是由于肿瘤细胞分泌的细胞因子 (IL-10,IL-8)、生长因子 (VEGF)、信号分子 (STAT3) 等影响了 DCs 的分化、成熟、功能[7]。
▐ T 细胞活化缺失
T 细胞的活化缺失,其主要原因是因为 APCs 的激活或者缺乏共刺激[8][9]。有研究证据表明缺乏或传递 FRP1 变体的 DCs 在抗原呈递和 T 细胞激活失败时,会导致乳腺癌及结直肠癌患者的肿瘤免疫反应,降低总体生存率[8]。
▐ T 细胞向肿瘤的运输受损
T 细胞向肿瘤的运输受损,主要由以下原因造成:免疫抑制肿瘤周围基质和肿瘤细胞改变对 CD8+ T 细胞的排斥作用,最近一项研究就将肿瘤基质细胞与肿瘤免疫中的 PD-L1 应答联系了起来[9]。而肿瘤细胞中 PTEN 的缺失可增加免疫抑制细胞因子的表达,从而导致 T 细胞介导的肿瘤杀伤被抑制,并减少 T 细胞向肿瘤运输[10]。
T 细胞向肿瘤的运输受损的另一个原因是趋化因子及细胞因子的影响,炎症环境下分泌的趋化因子可以使未成熟的 DCs 细胞向肿瘤聚集,并且由 DCs 细胞产生的趋化因子 CXCL16 及其受体 CXCR6 与 CD4+ 和 CD8+ T 细胞募集增有关,因此趋化因子的缺乏可能会导致 T 细胞的募集受损[11][12]。细胞因子则是 DCs 细胞产生活性的必需品,如由 DCs 产生的I型干扰素 (IFN-I) 可以以自分泌方式产生具有活性的 DC1s[13]。

针对冷肿瘤形成的机制,使冷肿瘤转变为热肿瘤的方法有:
① 使用可以诱导表观遗传的药物如 DNA 甲基转移酶抑制剂 DNMTi,来增强肿瘤抗原的表达[14]。
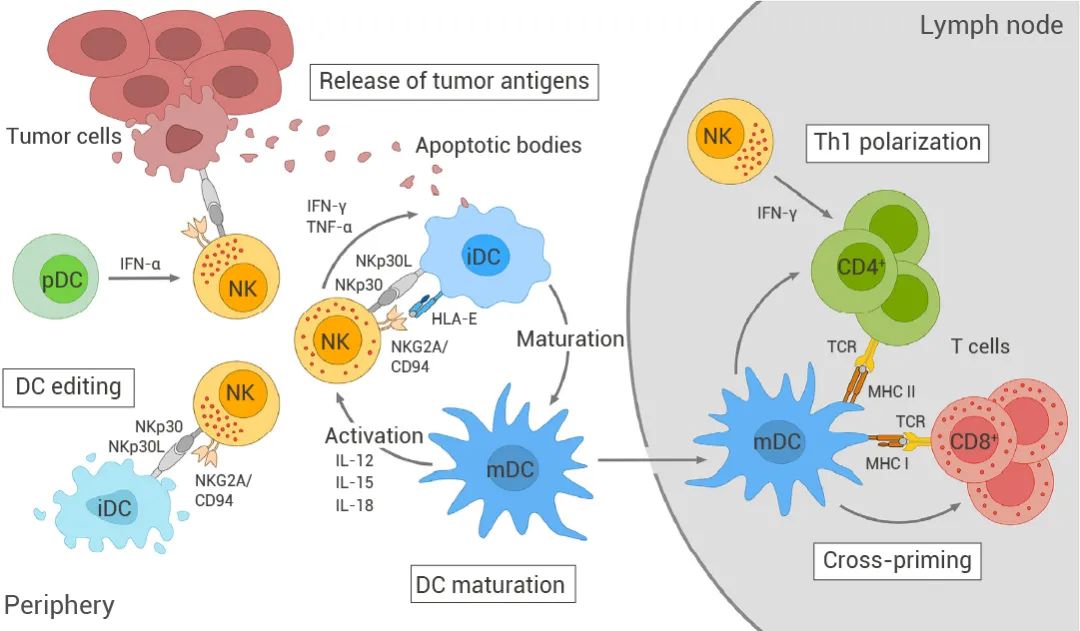
② 利用 NK 治疗法,如离体活化的自然杀伤细胞 (NK) 来改善肿瘤抗原缺乏或抗原呈递机制不足 (如图 3 所示)[15]。
③ 对于 T 细胞活化缺失则可以通过放化疗[16]、溶瘤病毒[17]、PRR 或 CD 40 激动剂[18][19]、肿瘤疫苗等方法来重新启动 T 细胞或与少量活化的 T 细胞产生协同作用[20]。
④ 若 T 细胞向肿瘤的运输受损,则可以将 TGF-β 抗体阻断剂和 TGF-β 受体拮抗剂与 PD-L1 抗体结合使用,共同诱导 T 细胞向肿瘤募集[21],也可以利用抗血管生成药物来增加T细胞在肿瘤细胞的浸润。
⑤ 开发将特定肿瘤抗原抗体片段与修饰细胞因子相结合免疫细胞因子,来特异性激活肿瘤内的免疫系统并减少全身副作用[22]。图 3. NK-DC 细胞互相干扰[15] 。 NK 细胞被靶肿瘤细胞或细胞因子激活后,会产生 IFN-γ 和肿瘤坏死因子 (TNF)-α,从而促进 DC 成熟。DC 的成熟也强烈依赖于 NK 细胞上激活受体(例如 NKp30 和 NKG2D)的参与。成熟的 DC (mDC) 反过来会产生白细胞介素 (IL)-12、IL-15 和 IL-18,增强 NK 细胞的细胞毒性和 IFN-γ 分泌。NK细胞还可以通过激活 NKp30 和抑制性杀伤细胞免疫球蛋白样受体以及 NKG2A/CD94 来区分未成熟(iDC)和 mDC 并消除未成熟 DC(iDC),从而维持 mDC 群体的质量(DC editing)。浆细胞样 DC (pDC) 分泌的 IFN-α 可以进一步增强 NK 细胞的细胞毒性。NK 诱导的肿瘤细胞裂解提供抗原,该抗原可以被 DC 吸收用于抗原呈递。一旦成熟,负载抗原的 mDC 将迁移到肿瘤引流淋巴结,将肿瘤抗原交叉呈递给初始 T 细胞,并诱导其分化为肿瘤特异性 CD8+ 细胞毒性 T 细胞和 CD4+ T 辅助 1 (Th1) 细胞。
以上的治疗方法中溶瘤病毒疗法以及肿瘤疫苗是被认为具有强大抗癌活性的新兴疗法。
 :溶瘤病毒疗法 :溶瘤病毒疗法
溶瘤病毒疗法不仅能够选择性的使肿瘤溶解外,而且通过溶瘤病毒裂解肿瘤细胞而诱导释放的 TAA、PAMP、DAMP 等可以激活体内的先天性和适应性免疫反应,改变肿瘤的免疫微环境使冷肿瘤变为热肿瘤[23]。在实际的应用中,T-VEC 就被证实是可以有效治疗黑色素瘤的溶瘤病毒[24]。临床上联合应用帕博利珠单抗能增加黑色素瘤患者的 CD8+ 细胞浸润及活化[25]。
:肿瘤疫苗
肿瘤疫苗则可以扩大特异性 T 细胞的数量,增加 T 细胞向肿瘤区域的运输[26]。在一项针对晚期恶性黑色素瘤、NSCLC 和膀胱癌患者的 Ib 临床试验中,个体化新抗原疫苗 NEO-PV-01 与 nivolumab 的联合就显著延长了无进展生存期,并观察到特异性T细胞向肿瘤区域的运输及浸润[27]。但无论是哪种治疗,最后都离不开肿瘤机制的探索,每一种机制的发现都会为我们的肿瘤治疗带来指导性的意义。
▐ MedChemExpressMCE 可提供 20,000+ 个用于肿瘤研究的相关产品及试剂,其中也包括肿瘤免疫微环境相关产品。

BIO8898
BIO8898 是一种有效的 CD40-CD154 抑制剂。BIO8898 抑制可溶性 CD40L 与 CD40-Ig 的结合, IC50 值为 25 μM。BIO8898 抑制 CD40L 诱导的细胞凋亡。 | Mitazalimab
Mitazalimab (ADC-1013; JNJ-64457107) 是 FcγR 依赖性 CD40 激动剂,具有肿瘤导向活性。Mitazalimab 激活抗原呈递细胞,例如 树突状细胞 (DC),以启动肿瘤反应性 T 细胞。因此,Mitazalimab 诱导肿瘤特异性 T 细胞浸润并杀死肿瘤。Mitazalimab 可重塑肿瘤浸润性骨髓微环境。 | TGFβ1-IN-1
TGFβ1-IN-1 (compound 42) 是一种有效的、具有口服活性的 TGF-β1 抑制剂。TGFβ1-IN-1 可以抑制 TGF-β1 诱导的纤维化标志物(α-SMA 和纤连蛋白)的上调,可用于肝纤维化疾病研究。 | SRI-011381 hydrochloride
SRI-011381 hydrochloride 是一种有具有口服生物活性的 TGF-β 信号通路的激活剂,具有神经保护作用。 | PD-1-IN-18
PD-1-IN-18 是 PD1 信号通路抑制剂,是一种免疫调节剂。 | CL845
CL845 是 STING 激动剂 CL656 (HY-112878) 的类似物。CL845 可用于合成靶向 STING(干扰素基因刺激物)的可结合 PRR 配体。CL845 可用于癌症、免疫系统疾病或感染的研究。 |
MCE的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。

[1] Yuan-Tong Liu, et al. Turning cold tumors into hot tumors by improving T-cell infiltration. Theranostics. 2021 Mar 11;11(11):5365-5386.
[2] Tumeh PC, et al. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature. 2014;515(7528):568–571.
[3] Mi Y, et al. The emerging role of myeloid-derived suppressor cells in the glioma immune suppressive microenvironment. Front Immunol. 2020;11:737.
[4] Bonaventura P, et al. Cold Tumors: A Therapeutic Challenge for Immunotherapy. Front Immunol. 2019 Feb 8;10:168.
[5] Wallich R, et al. Abrogation of metastatic properties of tumour cells by de novo expression of H-2K antigens following H-2 gene transfection. Nature. 1985;315(6017):301–305.
[6] Peng W, Chen JQ, Liu C, et al. Loss of PTEN Promotes Resistance to T Cell-Mediated Immunotherapy. Cancer Discov. 2016;6(2):202-216.
[7] Zhu S, et al. Tumor microenvironment-related dendritic cell deficiency: a target to enhance tumor immunotherapy. Pharmacol Res. 2020 Sep;159:104980.
[8] Vacchelli E, et al.. Chemotherapy-induced antitumor immunity requires formyl peptide receptor 1. Science (2015) 350:972–8.
[9] Mariathasan S, et al. TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature. 2018;554(7693):544-548.
[10] Toso A, et al. Enhancing chemotherapy efficacy in Pten-deficient prostate tumors by activating the senescence-associated antitumor immunity. Cell Rep. (2014) 9:75–89.
[11] Randolph GJ, et al. Migration of dendritic cell subsets and their precursors. Annu Rev Immunol. 2008;26:293-316.
[12] Hojo S, et al. High-level expression of chemokine CXCL16 by tumor cells correlates with a good prognosis and increased tumor-infiltrating lymphocytes in colorectal cancer. Cancer Res. 2007;67(10):4725-4731.
[13] Montoya M, et al. Type I interferons produced by dendritic cells promote their phenotypic and functional activation. Blood (2002) 99:3263–3271.
[14] Srivastava P, et al. Immunomodulatory action of SGI-110, a hypomethylating agent, in acute myeloid leukemia cells. Leuk Res. (2014) 38:1332.
[15] Zhang C, et al. Chimeric antigen receptor-engineered NK-92 cells: an off-the-shelf cellular therapeutic for targeted elimination of cancer cells and induction of protective antitumor immunity. Front Immunol. (2017) 8:533.
[16] Golden EB, et al. Radiotherapy and immunogenic cell death. Semin Radiat Oncol. 2015;25(1):11-17.
[17] Ribas A, et al. Oncolytic Virotherapy Promotes Intratumoral T Cell Infiltration and Improves Anti-PD-1 Immunotherapy [published correction appears in Cell. 2018 Aug 9;174(4):1031-1032]. Cell. 2017;170(6):1109-1119.e10.
[18] Bevers RFM, et al. Role of urothelial cells in BCG immunotherapy for superficial bladder cancer. Br J Cancer (2004) 91:607–12.
[19] Khong A, et al. The use of agonistic anti-CD40 therapy in treatments for cancer. Int Rev Immunol. (2012) 31:246–66.
[20] Ott PA, et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature (2017) 547:217–221.
[21] Shrimali RK, et al. Antiangiogenic agents can increase lymphocyte infiltration into tumor and enhance the effectiveness of adoptive immunotherapy of cancer. Cancer Res. 2010;70(15):6171-6180.
[22] Klein C, et al. Cergutuzumab amunaleukin (CEA-IL2v), a CEA-targeted IL-2 variant-based immunocytokine for combination cancer immunotherapy: overcoming limitations of aldesleukin and conventional IL-2-based immunocytokines.
Oncoimmunology (2017) 6:e1277306.
[23] Russell L, et al. Oncolytic Viruses: Priming Time for Cancer Immunotherapy. Biodrugs. 2019;33:485-501
[24] Andtbacka R, et al. Talimogene Laherparepvec Improves Durable Response Rate in Patients With Advanced Melanoma. J Clin Oncol. 2015;33:2780-8
[25] Ribas A, et al. Oncolytic Virotherapy Promotes Intratumoral T Cell Infiltration and Improves Anti-PD-1 Immunotherapy. Cell. 2017;170:1109-19.e10
[26] van der Burg SH, et al. Vaccines for established cancer: overcoming the challenges posed by immune evasion. Nat Rev Cancer. 2016;16:219-33.
[27]Ott P, Hu-Lieskovan S, Chmielowski B, et al. A Phase Ib Trial of Personalized Neoantigen Therapy Plus Anti-PD-1 in Patients with Advanced Melanoma, Non-small Cell Lung Cancer, or Bladder Cancer. Cell. 2020;183:347-62.e24
|
|


 |手机版|蒲公英|ouryao|蒲公英
( 京ICP备14042168号-1 ) 增值电信业务经营许可证编号:京B2-20243455 互联网药品信息服务资格证书编号:(京)-非经营性-2024-0033
|手机版|蒲公英|ouryao|蒲公英
( 京ICP备14042168号-1 ) 增值电信业务经营许可证编号:京B2-20243455 互联网药品信息服务资格证书编号:(京)-非经营性-2024-0033





 发表于 2024-2-1 16:22:42
发表于 2024-2-1 16:22:42


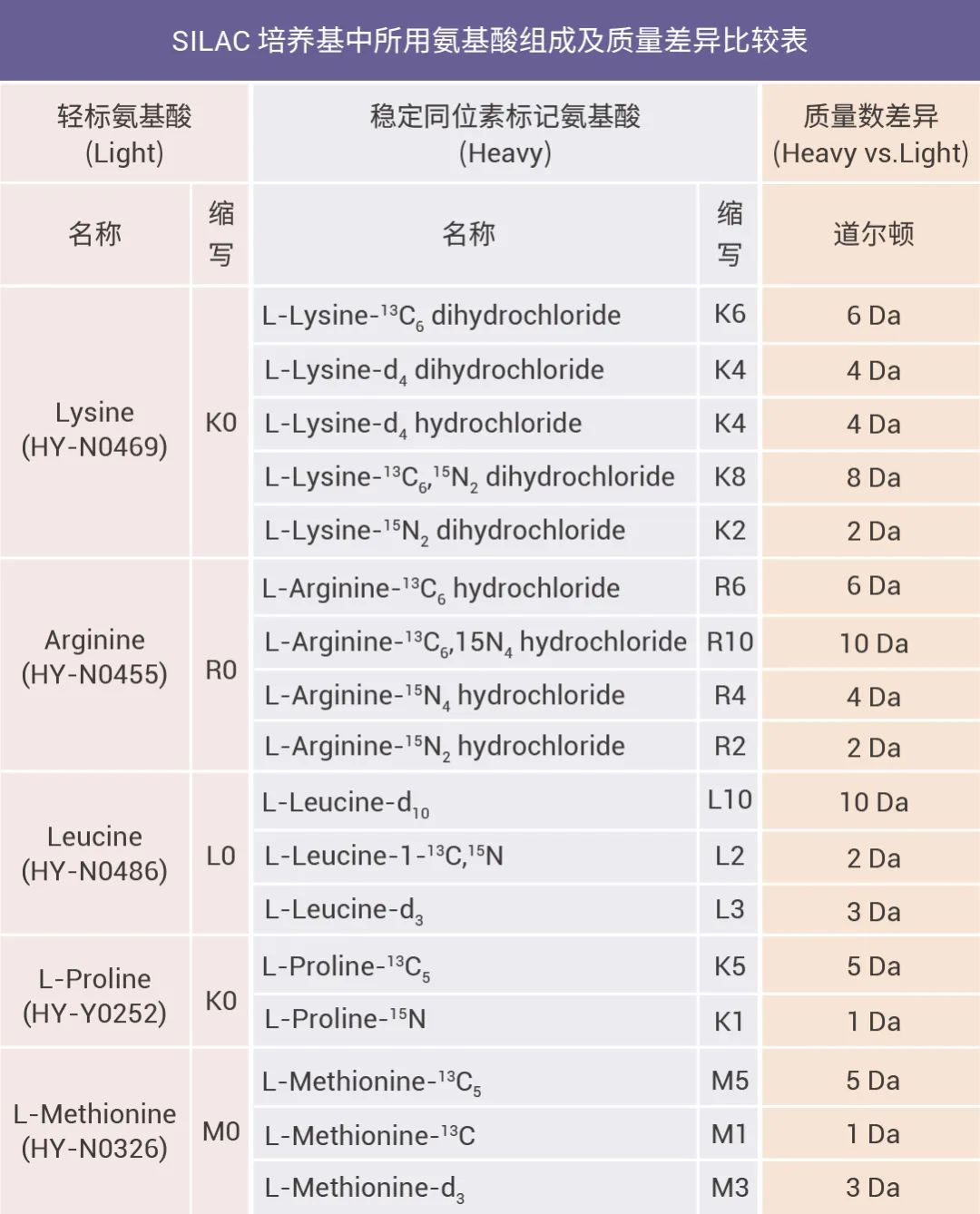

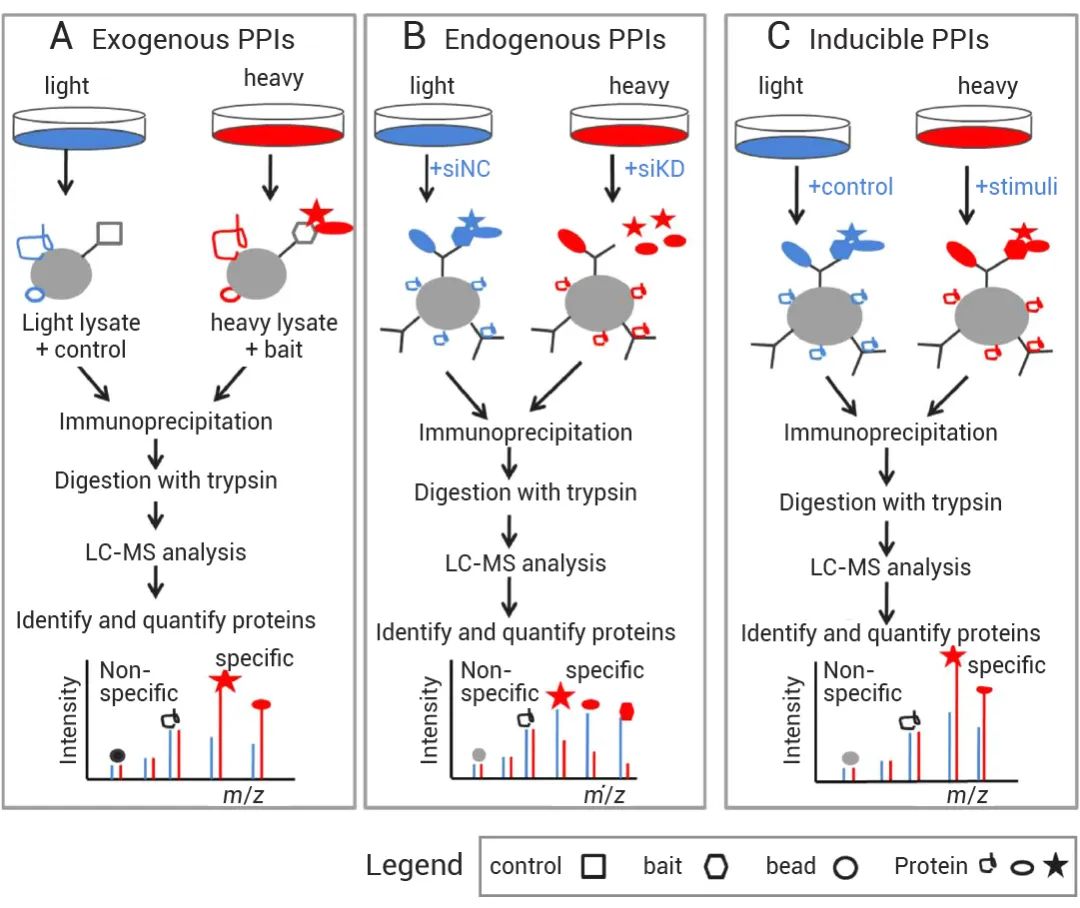


 置顶卡
置顶卡 变色卡
变色卡 类器官是由胚胎干细胞 (ESCs) 或诱导多能干细胞 (iPSCs) 衍生而来的体外 3D 细胞簇,与起源组织具有相似的组织学特征和功能。相比于传统 2D 培养,类器官更具生理相关性,已被广泛用于疾病研究和药物开发。
类器官是由胚胎干细胞 (ESCs) 或诱导多能干细胞 (iPSCs) 衍生而来的体外 3D 细胞簇,与起源组织具有相似的组织学特征和功能。相比于传统 2D 培养,类器官更具生理相关性,已被广泛用于疾病研究和药物开发。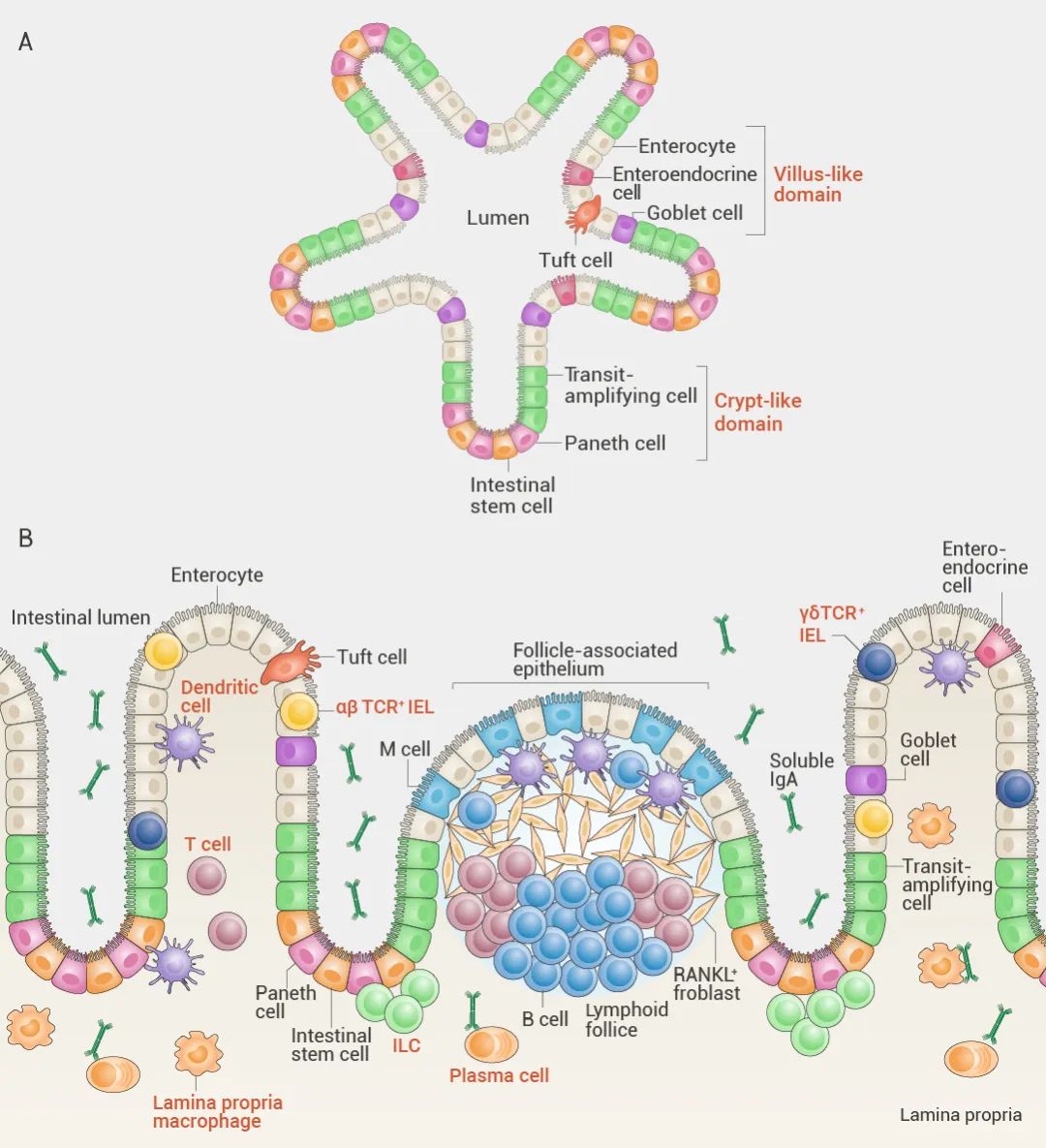

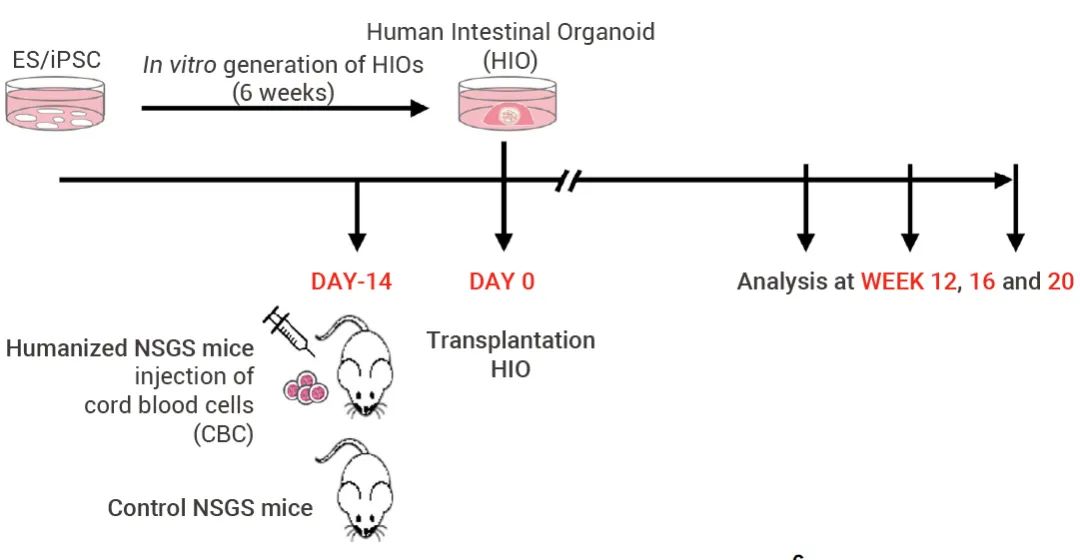
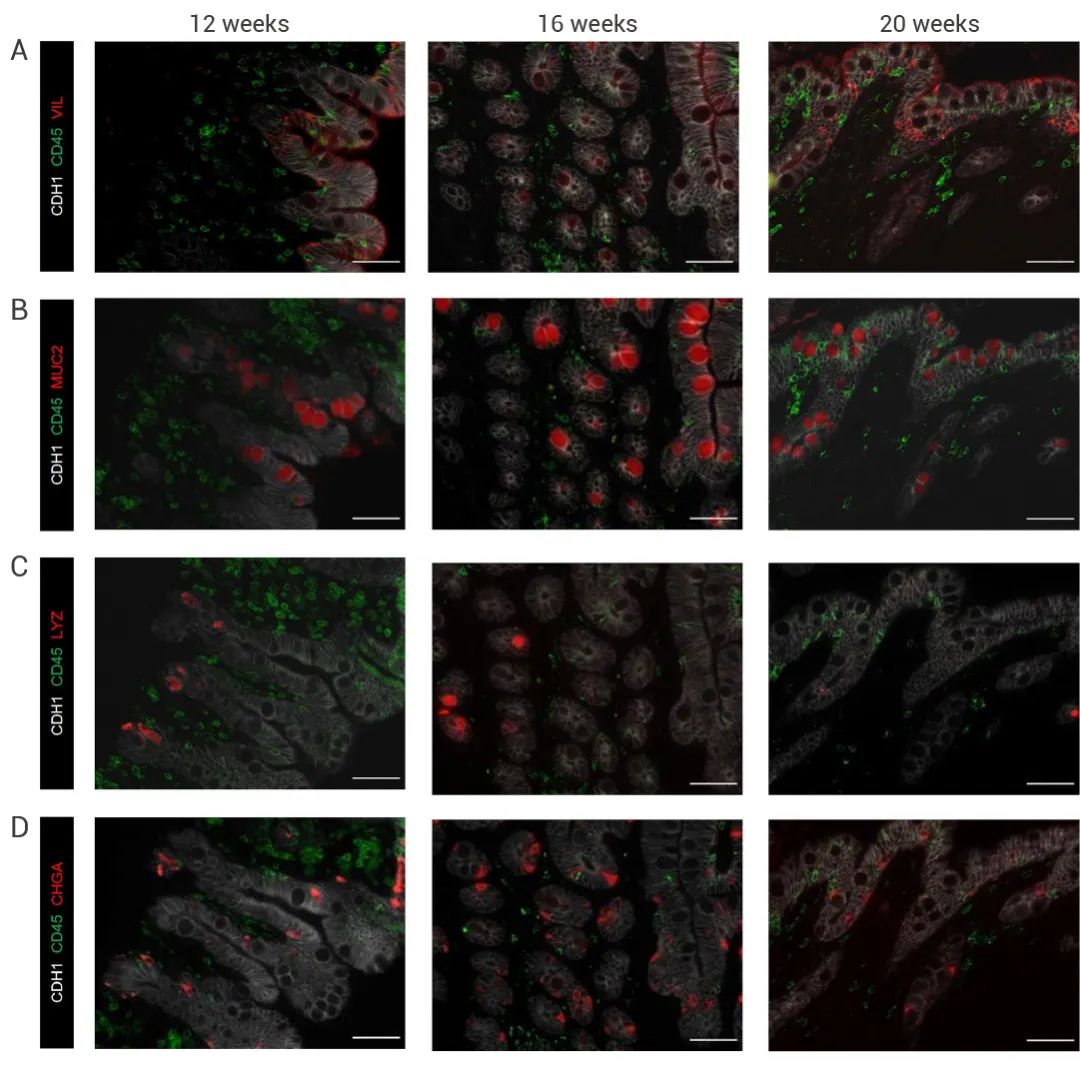
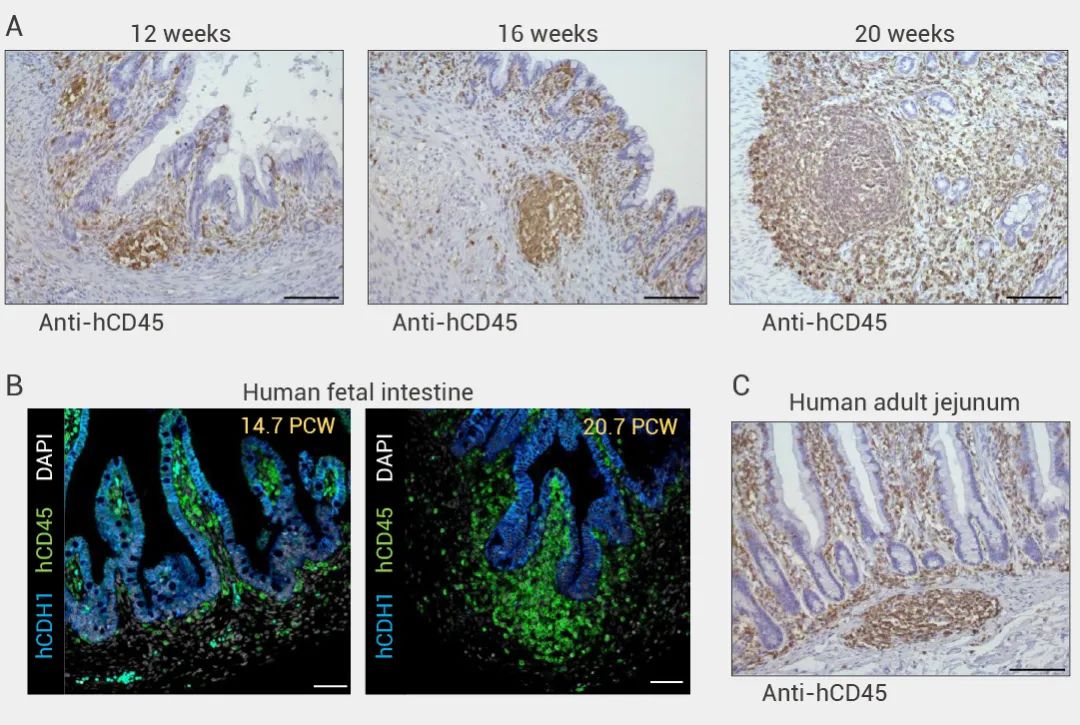
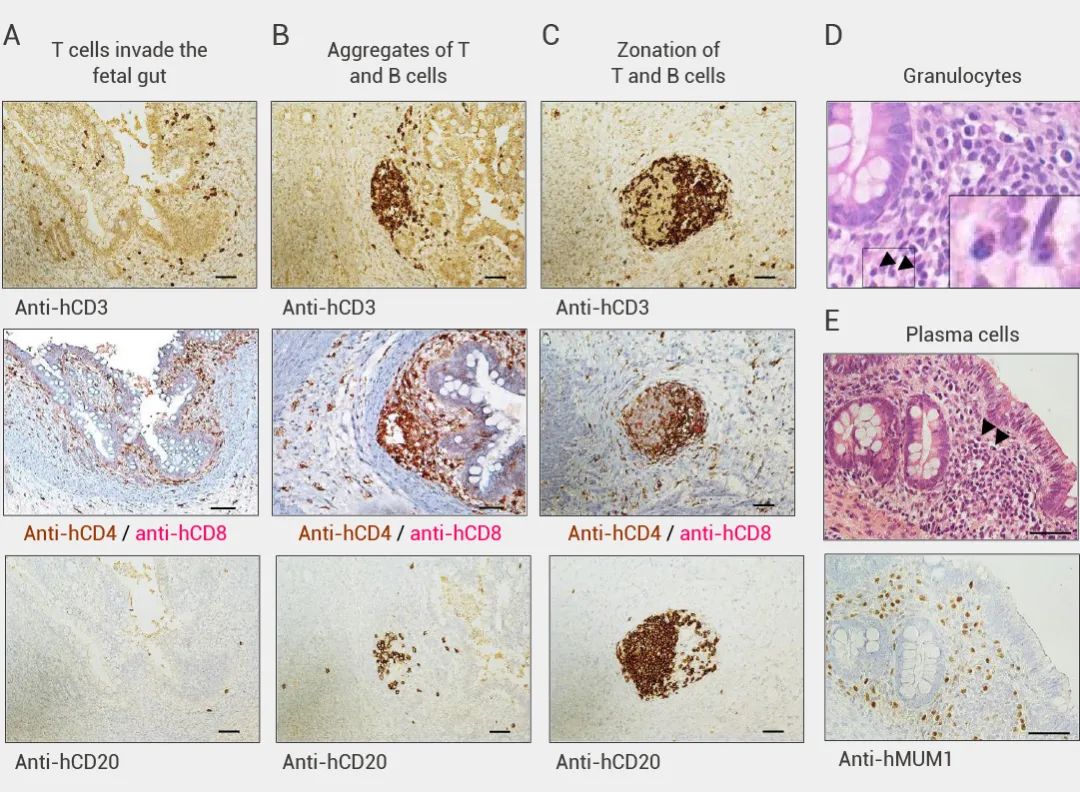
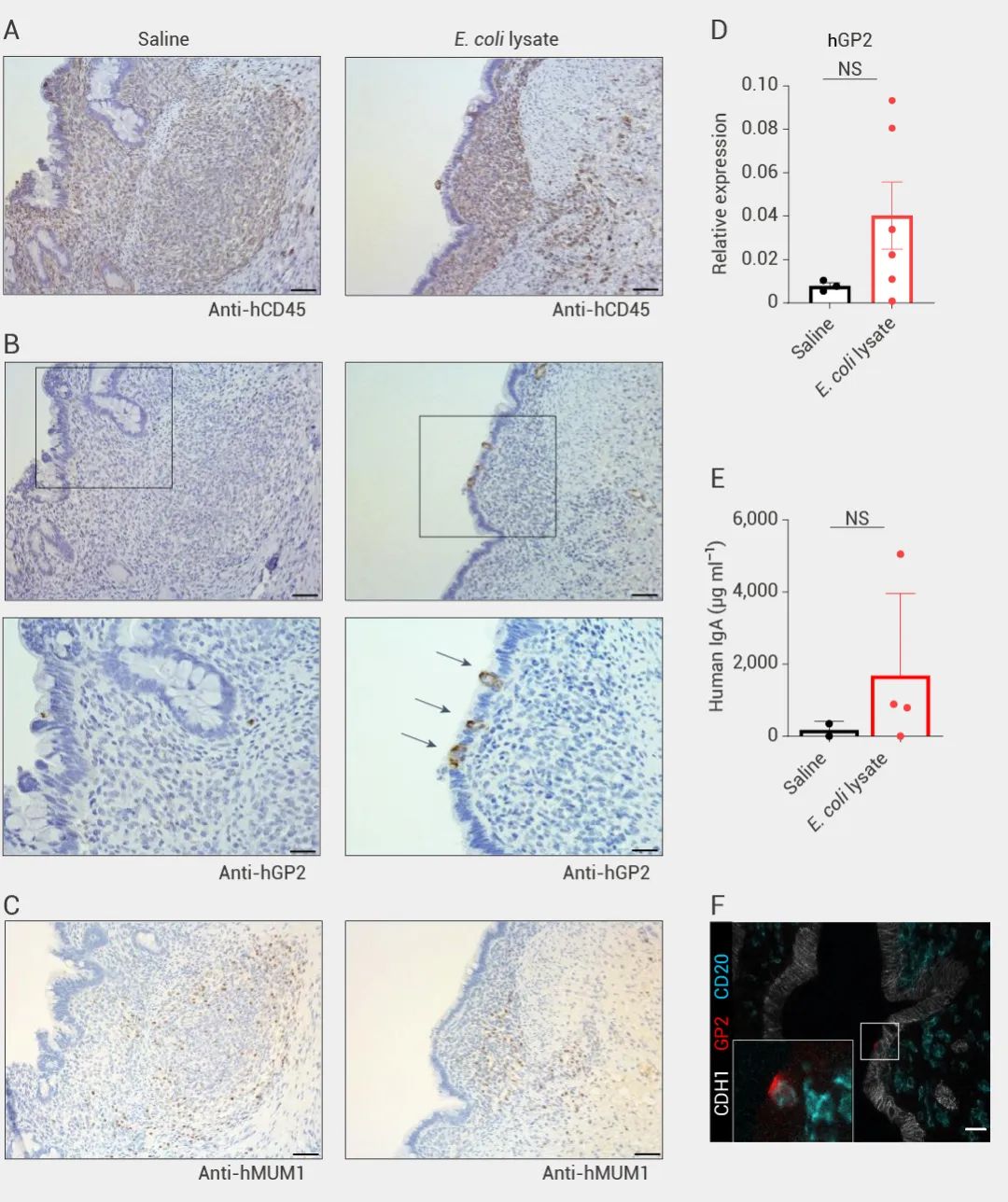
 本期小 M 为大家介绍了首例肠道“二代”类器官的构建与验证。该研究一定程度上弥补了免疫相关研究的缺陷,为研究肠道发育、相关疾病和药物发现的研究提供了更切实际的的模型。小 M 也相信这项研究也将为其他类器官领域的科学家们和同学们提供新思路~
本期小 M 为大家介绍了首例肠道“二代”类器官的构建与验证。该研究一定程度上弥补了免疫相关研究的缺陷,为研究肠道发育、相关疾病和药物发现的研究提供了更切实际的的模型。小 M 也相信这项研究也将为其他类器官领域的科学家们和同学们提供新思路~

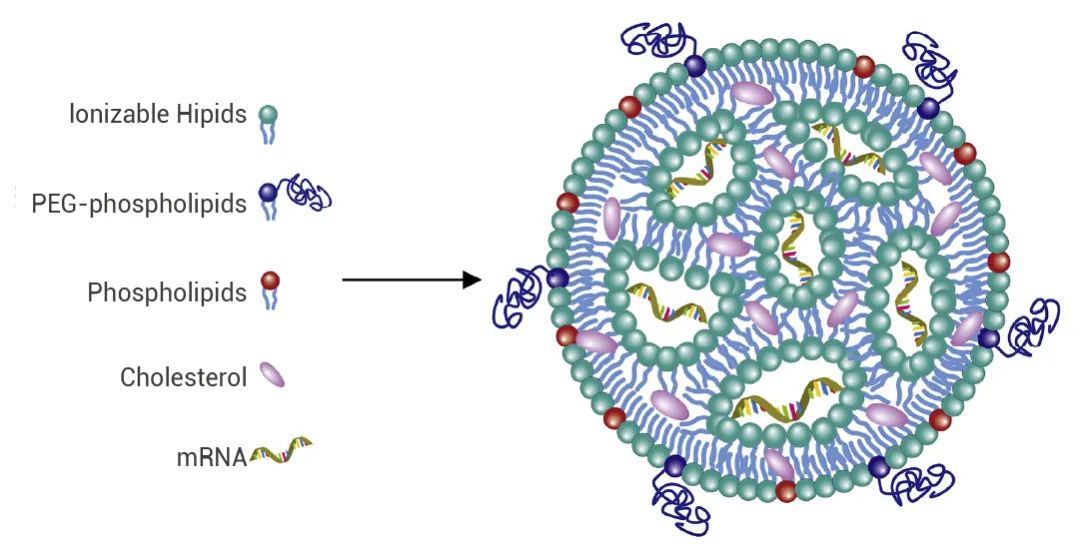


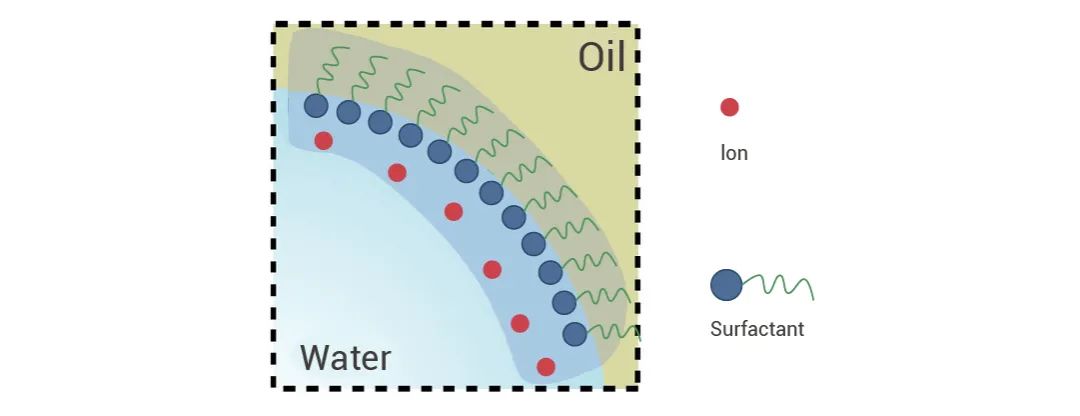
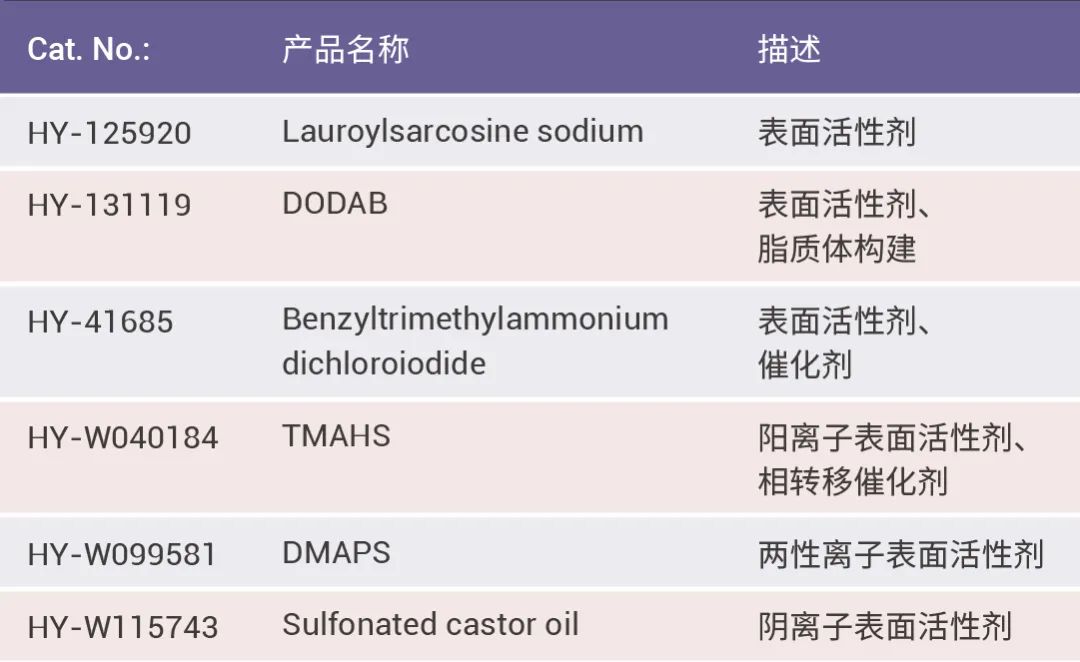
 一部分表面活性剂家族成员在助溶剂队伍里发光发热,看看有没有老朋友~
一部分表面活性剂家族成员在助溶剂队伍里发光发热,看看有没有老朋友~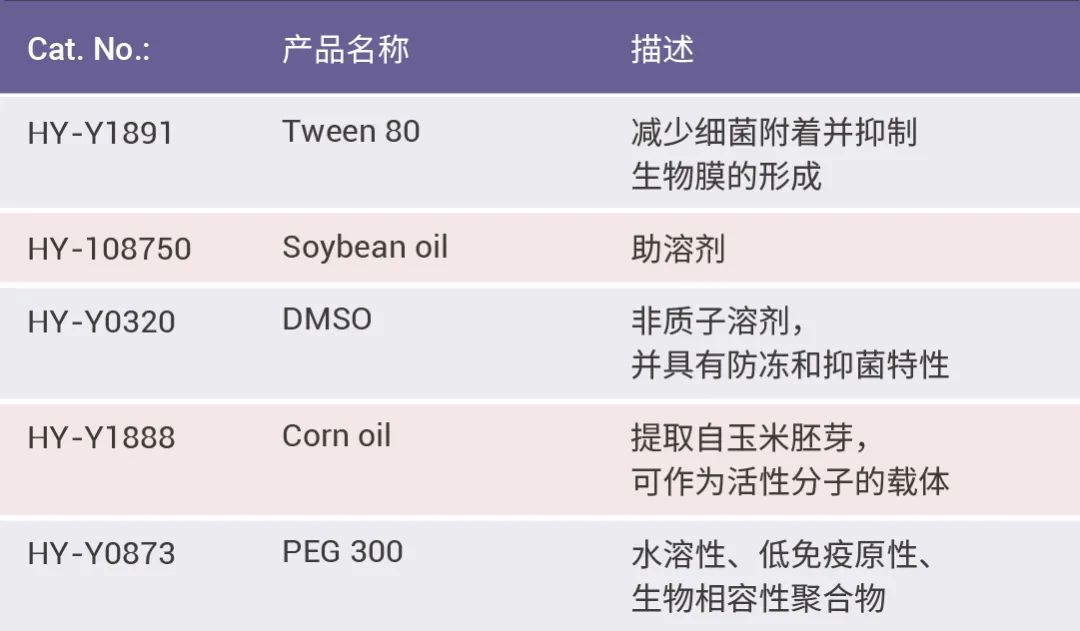


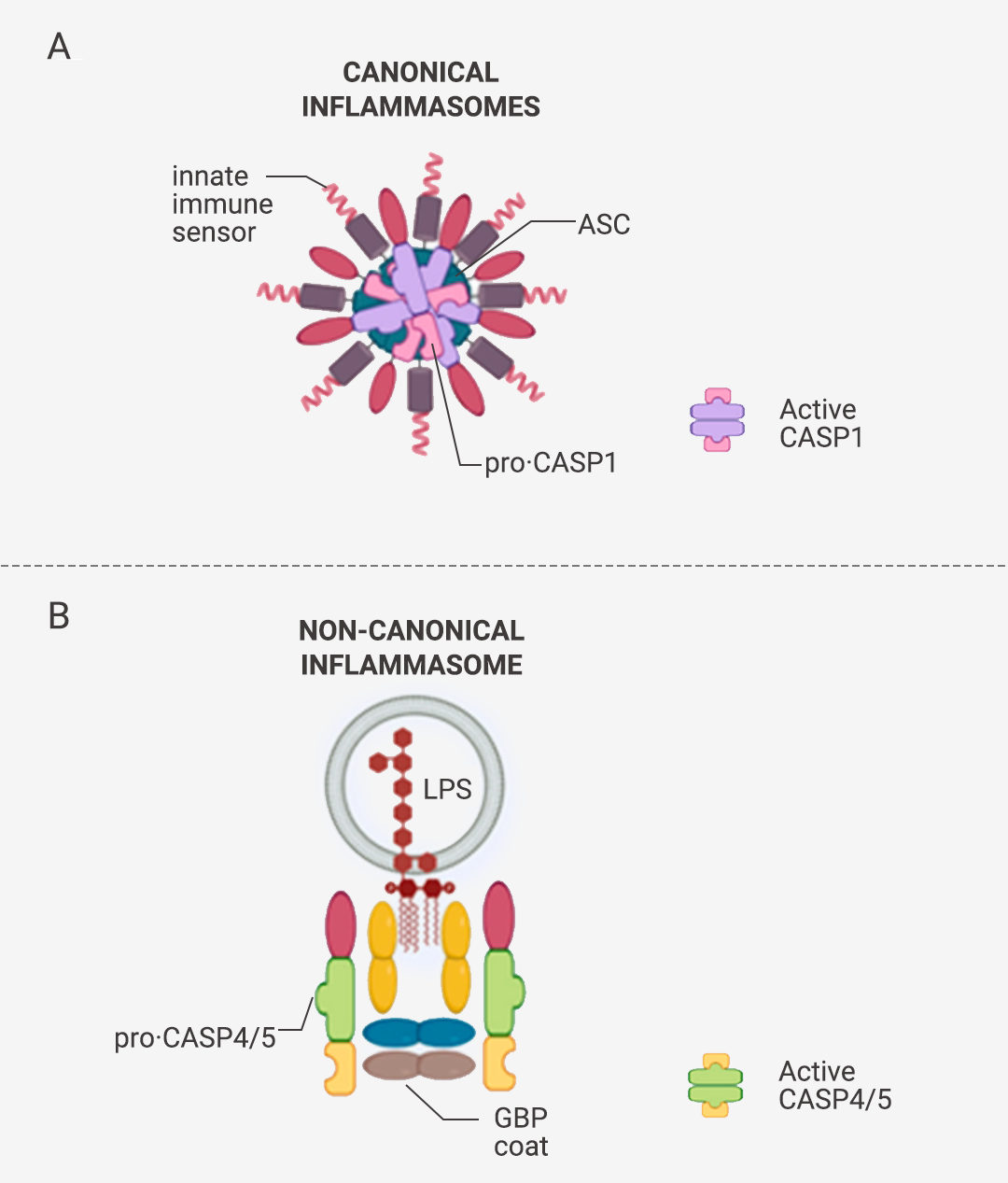
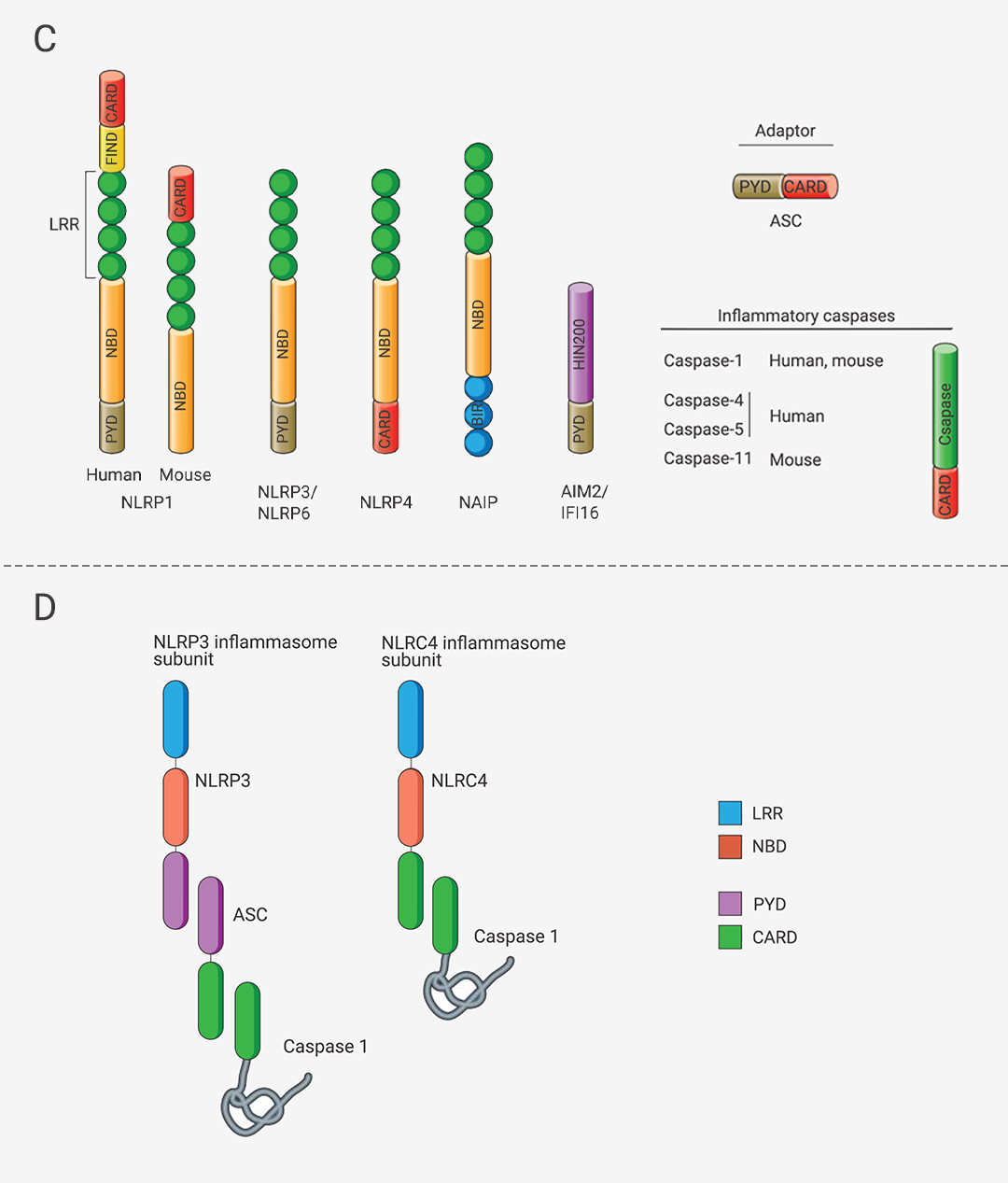
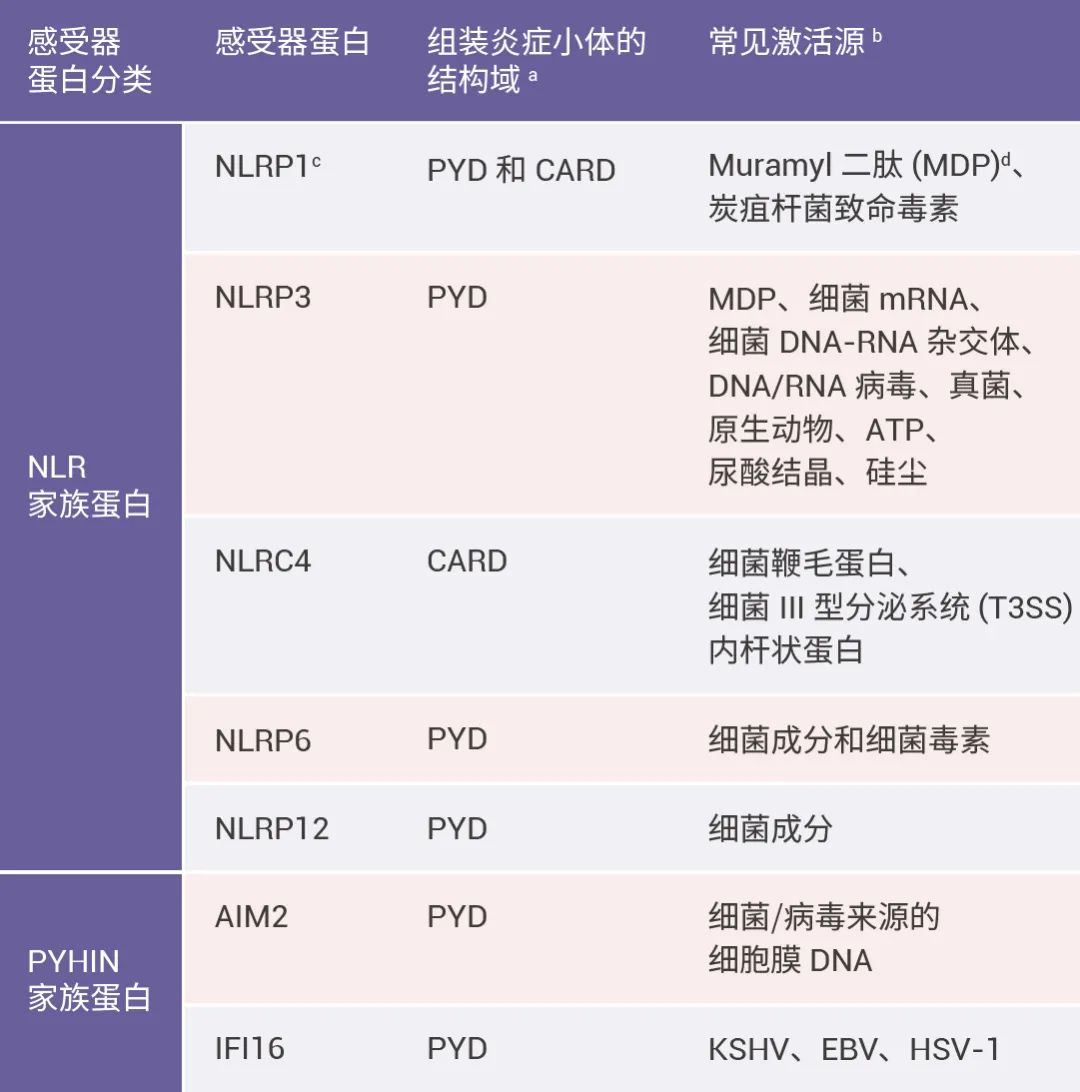

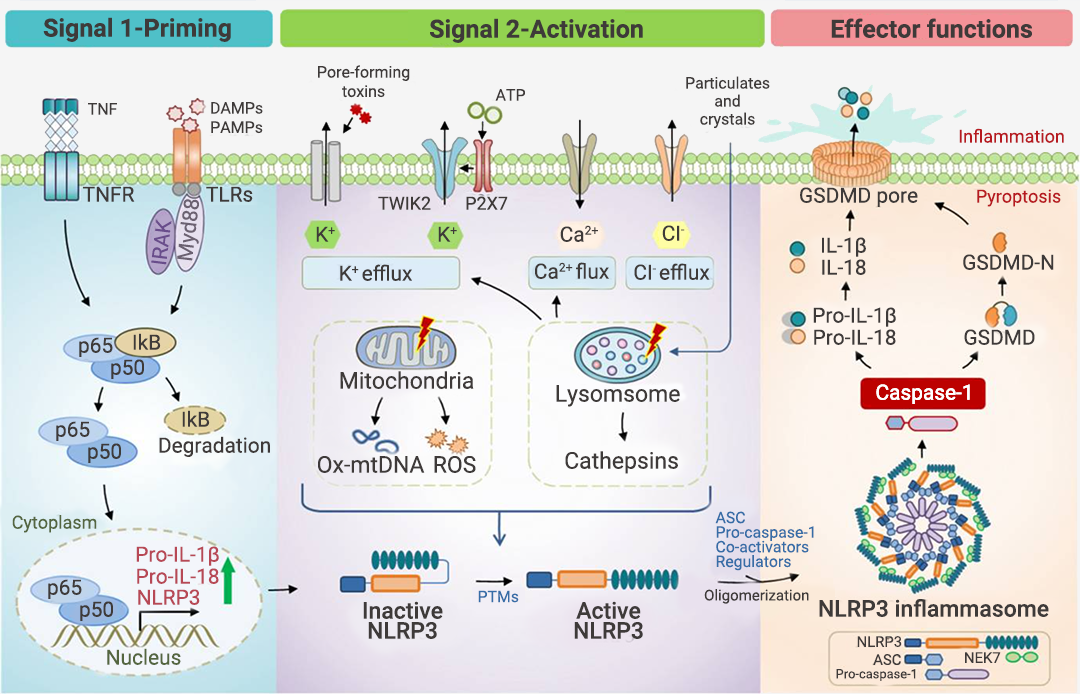
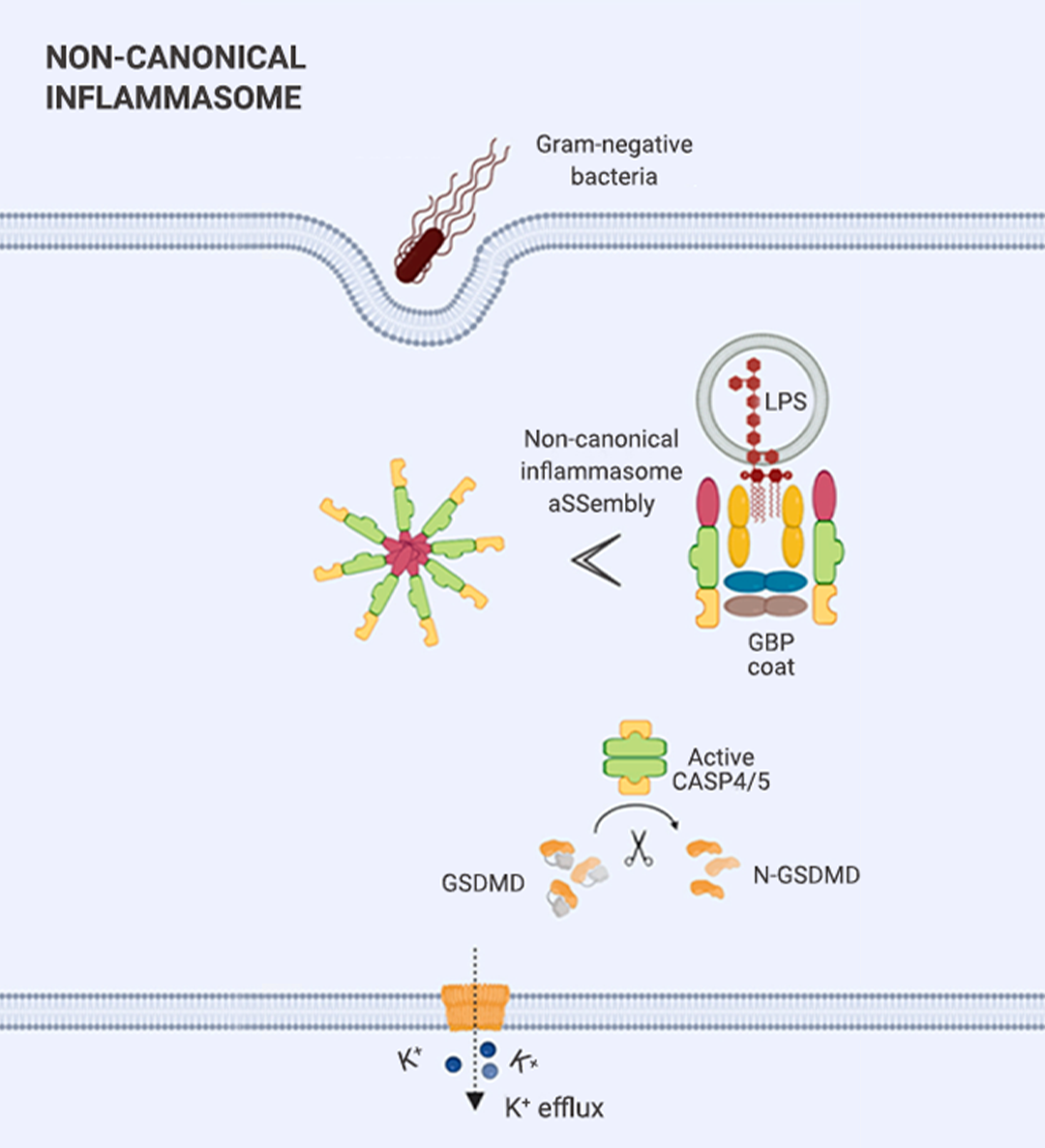
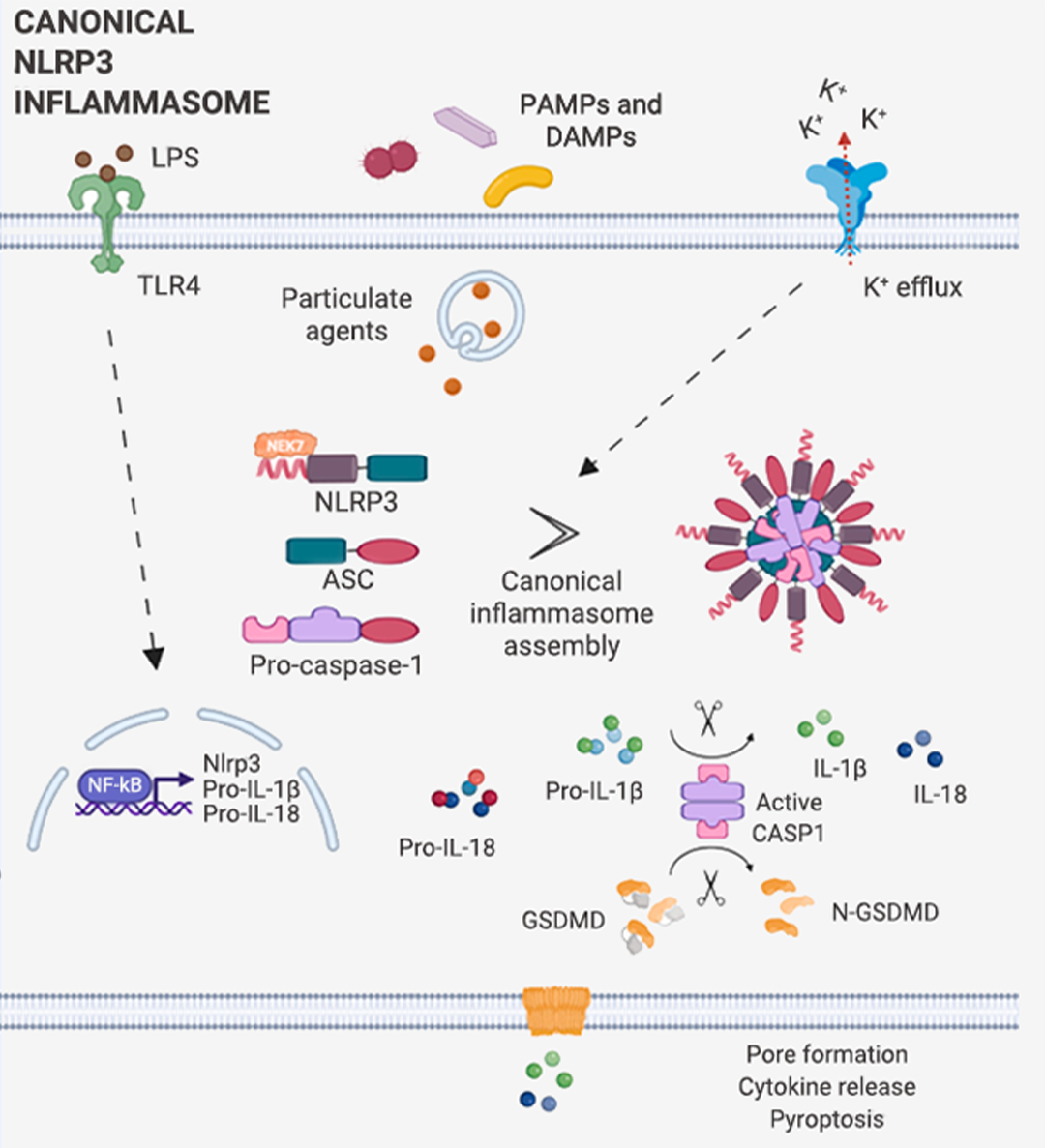
 炎性小体介导的炎症与多种人类疾病有很大关系,在急性炎症中,炎症小体的激活有助于去除死细胞并启动组织修复。然而,在慢性炎症中,炎症小体的持续激活是有害的,因为它会损伤组织。
炎性小体介导的炎症与多种人类疾病有很大关系,在急性炎症中,炎症小体的激活有助于去除死细胞并启动组织修复。然而,在慢性炎症中,炎症小体的持续激活是有害的,因为它会损伤组织。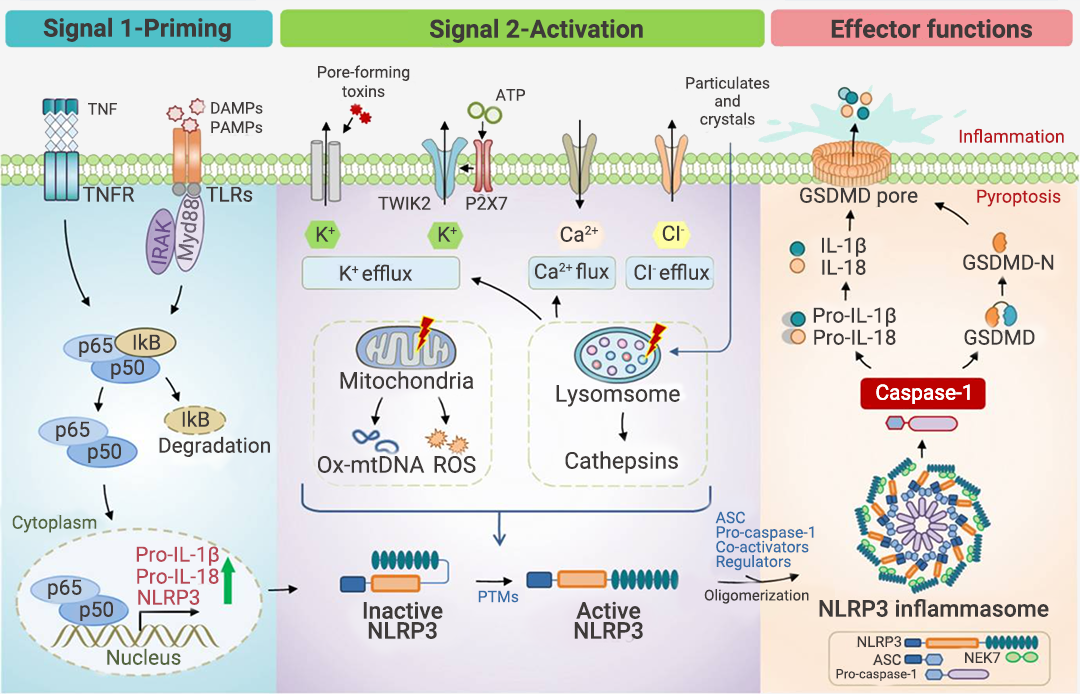

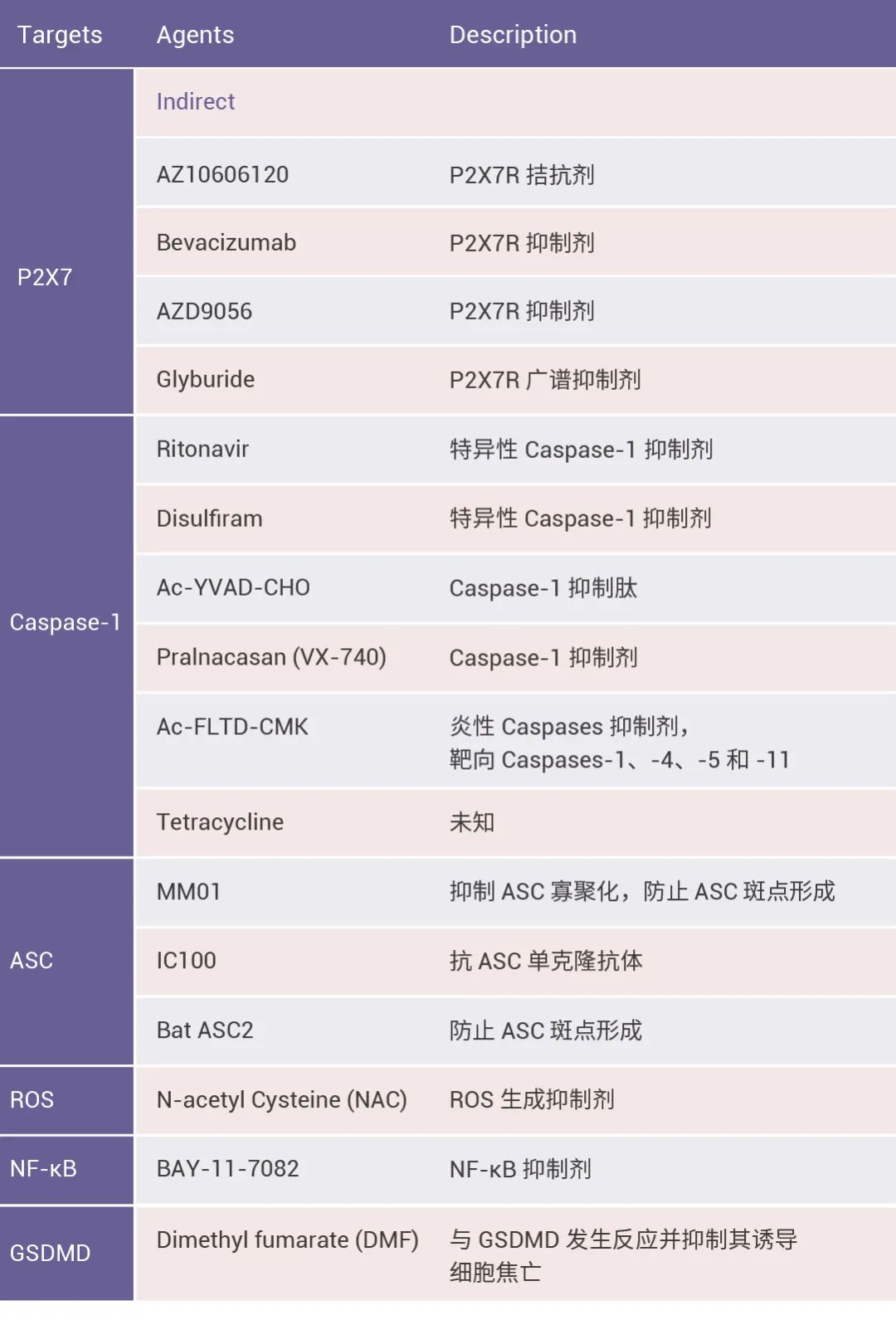
 本篇推文为大家详细介绍了炎症小体的分类 (经典和非经典炎症小体)、炎症小体的活化通路、炎症小体的组装方式、以及炎症小体涉及的人类疾病和 NLRP3 炎症小体靶向试剂的研发进展。这些现有成果是 20 多年来众多研究者一步一脚印所摘取的,但我们所揭露的也只是炎症小体的冰山一角,更广阔的未知犹待你我探索!
本篇推文为大家详细介绍了炎症小体的分类 (经典和非经典炎症小体)、炎症小体的活化通路、炎症小体的组装方式、以及炎症小体涉及的人类疾病和 NLRP3 炎症小体靶向试剂的研发进展。这些现有成果是 20 多年来众多研究者一步一脚印所摘取的,但我们所揭露的也只是炎症小体的冰山一角,更广阔的未知犹待你我探索! 
 [1] Martinon F, et al. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002 Aug;10(2):417-26.
[1] Martinon F, et al. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell. 2002 Aug;10(2):417-26.
